
Քարաքոսեր
 Պատկեր Հեկկելի «Kunstformen der Natur» գրքից, 1904 | |||
| Անվանում | |||
|---|---|---|---|
| Քարաքոսեր | |||
| Անվանման կարգավիճակ | |||
| Հնացած կարգաբանական | |||
| Լատիներեն անվանում | |||
| Lichenes | |||
| Մայր տաքսոն | |||
| Ենթաթագավորություն Ցածրակարգ բույսեր (Thallophyta) | |||
| |||
Քարաքոսեր (լատին․՝ Lichenes; laɪkəns, նաև lɪtʃəns)[1]), բարդ օրգանիզմներ են՝ կազմված սնկից (միկոբիոնտ) և ֆոտոսինթեզող օրգանիզմից (ֆոտոբիոնտ), որոնք աճում են միասին։ Նրանց միջև գոյություն ունեն սիմբիոտիկ հարաբերություններ։ Ֆոտոբիոնտը սովորաբար կանաչ ջրիմուռ է (հիմնականում Trebouxia ցեղը) կամ ցիանոբակտերիա (հիմնականում Nostoc ցեղը)[2]։ Քարաքոսերի մորֆոլոգիան, ֆիզիոլոգիան և կենսաքիմիան բավականին տարբերվում են առանձին վերցրած բաղադրիչների նույն հատկություններից։ Քարաքոսերի կարելի է հանդիպել երկրագնդի մի շարք էքստրեմալ միջավայրերում ՝ արկտիկական տունդրաներում, չոր անապատներում, ժայռոտ ծովափերում։ Նրանք հաճախ աճում են նաև խոնավ անտառներում՝ ապրելով ծառերի ճյուղերի և բների վրա (որպես էպիֆիտներ), մերկ ժայռերի վրա, պատերի, խաչքարերի վրա, ինչպես նաև հողի մակերեսին։ Քարաքոսերը լայնորեն տարածված են և հավանաբար երկարակյաց են[3]։ Բայց և այնպես շատ քարաքոսեր խոցելի են միջավայրի պայմանների փոփոխությունների նկատմամբ, ուստի հարմար գործիք են գիտնականների համար օդի աղտոտվածությունը, օզոնային շերտի քայքայումը և մետաղներով աղտոտումը գնահատելու համար[4][5]։ Քարաքոսերը օգտագործվել են նաև ներկերի և օծանելիքների ստացման համար, ինչպես նաև՝ ավանդական բժշկության մեջ։
Ընդհանուր տեղեկություններ[խմբագրել | խմբագրել կոդը]

Շատ քարաքոսերի մարմինը՝ թալոմը, տարբերվում է այն կազմող և առանձին աճող սնկի կամ ջրիմուռի մարմնից։ Սունկը շրջապատում է ջրիմուռի բջիջները քարաքոսերի համակեցություններին բնորոշ բարդ սնկային հյուսվածքով։ Քարաքոսերի շատ տեսակների մոտ սունկը ներթափանցում է ջրիմուռի բջիջների բջջապատի մեջ՝ առաջացնելով հաուստորիումներ, որոնք նման են պաթոգեն սնկերի մոտ առկա կառուցվածքներին։ Քարաքոսերը պոյկիլոհիդրիկ օրգանիզմեր են՝ կարող են գոյատևել ջրի չափազանց ցածր պարունակության պայմաններում։ Բջջաթաղանթների վերակազմավորումը, ինչին հաջորդում է ջրազրկման փուլը, տևում է ամենաքիչը մի քանի րոպե։
Ջրիմուռի կամ ցիանոբակտերիայի բջիջները կատարում են ֆոտոսինթեզ և բույսերի նման վերականգնում են մթնոլորտային ածխածնի երկօքսիդը օրգանական նյութերի և դրանով ապահովում իրենց և սնկի սնուցումը։ Քարաքոսի երկու բաղադրիչներն էլ ջուրը և հանքային նյութերը գլխավորապես ձեռք են բերում մթնոլորտից՝ անձրևի և փոշու միջոցով։ Սնկային բաղադրիչը պաշտպանում է ջրիմուռին ջուր կուտակելու միջոցով և ծառայում է որպես հանքային նյութերի պահեստ՝ որոշ դեպքերում ապահովելով ջրիմուռին սուբստրատից ձեռք բերված հանքանյութերով։ Եթե քարաքոսի կազմում առկա է ցիանոբակտերիա՝ որպես առաջնային բաղադրիչ կամ լրացուցիչ սիմբիոնտ (երեք մասից կազմված ջրիմուռների մոտ), ապա հնարավոր է մթնոլորտային ազոտի ֆիքսում։
Չնայած տարբեր ցիանոքարաքոսերում գտնված ցիանոբակտերիաների շտամները հաճախ իրար նման են, նրանք տարբերվում են ազատ ապրող նույն շտամներից։ Քարաքոսերում տեղի ունի իսկական սիմբիոզ։ Այն մեծացնում է երկու բաղադրիչների էկոլոգիական հնարավորությունները, բայց բնական միջավայրերում նրանց աճի և բազմացման համար պարտադիր չէ, քանի որ ջրիմուռ-սիմբիոնտներից շատերը կարող են ապրել ազատության մեջ։ Օրինակ՝ Trentepohlia ջրիմուռը առաջացնում է նարնջագույն պոպուլյացիաներ՝ ծառերի բների և համապատասխան ժայռերի մակերեսներին։ Քարաքոսերի համակեցություններում կարող է դիտվել մուտուալիզմ, կոմենսալիզմ և նույնիսկ մակաբուծություն՝ կախված այն կազմող տեսակներից։ Պարզվել է, որ ցիանոբակտերիաները լաբորատոր պայմաններում կարող են աճել ավելի արագ, քան քարաքոսի կազմում լինելու ժամանակ։
2012 թվականի ապրիլի 26-ին, գիտնականները հաղորդեցին, որ «Մարսի սիմուլյացիայի լաբորատորիա»-ում Մարս մոլորակի պայմանների սիմուլյացիայի ժամանակ՝ 34 օրվա ընթացքում, քարաքոսը գոյատևել է և ցույց է տվել նշանակալի արդյունքներ՝ ֆոտոսինթետիկ ակտիվության հարմարողականության հետ կապված[6][7]։
Պատմություն[խմբագրել | խմբագրել կոդը]
Չնայած քարաքոսերը վաղուց հայտնի են եղել որպես օրգանիզմներ, միայն 1867 թվականին շվեյցարացի գիտնական Սիմոն Շվենդեները առաջարկեց քարաքոսերի երկակի բնույթի վերաբերյալ իր տեսությունը[8], և քարաքոսերի համակեցությունների իրական բնույթը սկսեց աստիճանաբար բացահայտվել։ Շվենդեների հիպոթեզը, որն այդ ժամանակ չուներ փորձնական ապացույցներ նրա կողմից լուսային մանրադիտայով քարաքոսերի, սնկերի և ջրիմուռների անատոմիայի և զարգացման երկարատև ուսումնասիրությունների վրա էր հիմնված։ Այդ ժամանակվա հայտնի քարաքոսաբաններից շատերը, այդ թվում Ջեյմս Քրոմբին և Նիլանդերը ժխտում էին Շվենդեների հիպոթեզը՝ պնդելով, որ բոլոր կենդանի օրգանիզմները ավտոնոմ էին։ Այլ նշանավոր գիտնականներ, ինչպիսիք են Հենրիխ Անտոն դե Բարին, Ալբերտ Բեռնհարդ Ֆրանկը, Մելքիոր Թրեուբը և Հերման Հելռիգելը այդ վարկածը չէին բացառում։ Երկակի բնույթի մասին գաղափարները շուտով տարածվեցին նաև կենսաբանության այլ բնագավառներում։ Երբ վերջապես բացահայտվեցին պաթոգեն միկրոօրգանիզմների և նրանց տեր բջիջների միջև բարդ փոխազդեցությունները, Շվենդեների հիպոթեզը սկսեց ճանաչում ձեռք բերել։ Հետագայում Էուգեն Թոմասը տվեց քարաքոսերի երկակի բնույթի վերաբերյալ փորձնական ապացույցներ և տպագրեց իր արդյունքները 1939 թվականին[9]։
Սիմբիոնտներ[խմբագրել | խմբագրել կոդը]
Սնկի համար քարաքոսի կազմում սիմբիոնտի կարգավիճակով ապրելը շահավետ է սննդանյութեր ստանալու առումով և սնկերի տեսակների մոտավորապես 80%-ը ձեռք է բերել այսպիսի կենսակերպ։ Ամենատարածված լիխենիզացված սնկերը ասկոմիցետներն են։ Նրանց մոտավորապես 40%-ը առաջացնում է այսպիսի ասոցիացիաներ։ Այս լիխենիզացված սնկերի մի մասը հանդիպում է նաև որպես ազատ ապրող ձևեր՝ չլիխենիզացվող սնկերի կարգերում՝ որպես սապրոտրոֆներ կամ բույսերի մակաբույծներ (օրինակ՝ կարգ Leotiales, կարգ Dothideales և կարգ Pezizales), իսկ մյուս մասը միայն քարաքոսերի կազմում (օրինակ՝ կարգ Graphidales, կարգ Gyalectales, կարգ Peltigerales, կարգ Pertusariales և կարգ Teloschistales)։ Սակայն նույնիսկ միևնույն ցեղի կամ տեսակի սահմաններում կարող են հանդիպել և լիխենիզացված և չլիխենիզացված սնկեր։ Ընդհանուր առմամբ քարաքոսերի մոտավորապես 98%-ի միկոբիոնտը ասկոմիցետներն են։ Քարաքոսերի կազմում լ հաճախ հանդիպող սնկերի երկրորդ խումբը անկատար սնկերն են։ Համեմատաբար քիչ քանակությամբ հանդիպում են բազիդիոմիցետներ, որոնց թվում են Lichenomphalia, Multiclavula և Dictyonema ցեղերի ներկայացուցիչներ։
Քարաքոսերում հանդիպող ավտոտրոֆ սիմբիոնտները ֆոտոսինթետիկ ջրիմուռներն են։ Դրանք կարող են լինել ինչպես էուկարիոտ, այնպես էլ պրոկարիոտ օրգանիզմներ։ Հայտնաբերվել է ֆոտոսինթետիկ օրգանիզմների մոտավորապես 100 տեսակ, որոնք առաջացնում են ասոցիացիաներ քարաքոս առաջացնող սնկերի հետ։ Պրոկարիոտ օրգանիզմները ցիանոբակտերիաներն են, որոնց երբեմն կոչում են նաև կապտականաչ ջրիմուռներ։ Նրանք կազմում են հայտնի քարաքոսերի 8%-ի ֆոտոսինթետիկ սիմբիոնտը, ընդ որում ամենից հաճախ հանդիպող ցեղը Nostoc-ն է։ Սակայն քարաքոսերի գերակշռող մասը պարունակում է էուկարիոտ ավտոտրոֆներ, որոնցից են կանաչ (բաժին Chlorophyta) և դեղնականաչ ջրիմուռները (բաժին Xanthophyta)։ Քարաքոսերի 90%-ի մոտ հանդիպում են կանաչ ջրիմուռներ, և մասնավորապես Trebouxia ցեղը ամենից հաճախ հանդիպողներից է (քարաքոսերի 40%-ի մոտ)։ Երկրորդ տեղում է Trentepohlia ցեղը։ Այսպիսով, հայտնի է ջրիմուռների մոտավորապես 100 տեսակ, որոնք որպես ավտոտրոֆներ հանդիպում են քարաքոսերի կազմում։ Հավանաբար բոլոր այդ ջրիմուռներն էլ կարող են ազատ կերպով գոյատևել բնության մեջ՝ քարաքոսից անկախ[10]։
Որոշակի սնկերի և ջրիմուռների տեսակներ քարաքոսի կազմում իրար հետ միակ ձևով չեն կապված։ Օրինակ՝ մեկ սունկը կարող է քարաքոս առաջացնել տարբեր տեսակի ջրիմուռների հետ։ Որոշակի սնկային սիմբիոնտի կողմից տարբեր ջրիմուռների հետ ստեղծած քարաքոսերի թալոմները կլինեն իրար նման, երկրորդային մետաբոլիտները՝ նույնպես, այսպիսով, ցույց տալով, որ քարաքոսի արտաքին տեսքի ձևավորման հարցում որոշիչ դերը պատկանում է սնկին։ Ավելին՝ միևնույն ջրիմուռի տեսակը կարող է հանդիպել տարբեր տեսակի սնկերի հետ ասոցացված վիճակում։ Հայտնի են նաև այնպիսի քարաքոսեր, որոնցում մեկ սունկը միացած է երկու և երեք տեսակի ջրիմուռների հետ միևնույն ժամանակ։ Հազվադեպ հանդիպում է նաև հակառակը, երբ մի քանի սնկեր փոխազդում են իրար հետ և առաջացնում քարաքոս։
Ե´վ քարաքոսը, և´ սնկային բաղադրիչը ունեն միևնույն գիտական անվանումը և քարաքոսերը դասակարգելիս հիմնվում են սնկերի դասակարգման վրա։ Ջրիմուռը ունի իր գիտական անվանումը, որը կապ չունի քարաքոսի կամ սնկի անվանման հետ։
Մորֆոլոգիա և կառուցվածք[խմբագրել | խմբագրել կոդը]



Որոշ քարաքոսեր ունեն տերևների տեսք (տերևանման քարաքոսեր), որոշները ծածկում են սուբստրատը կեղևի նման (կեղևանման քարաքոսեր), մյուսները, օրինակ՝ Ramalina ցեղի ներկայացուցիչները, ունեն են թփի տեսք (թփանման քարաքոսեր), և կան նաև դոնդողանման քարաքոսեր, օրինակ՝ Collema ցեղի ներկայացուցիչները։
Չնայած քարաքոսի ձևը կանխորոշված է սնկային բաղադրիչի գենետիկական նյութով, ֆոտոբիոնտի հետ կապը անհրաժեշտ է համապատասխան ձևի ձևավորման համար։ Երբ լաբորատոր պայմաններում սունկը աճեցվում է առանց ֆոտոբիոնտի առկայության, այն ուղղակի զարգանում է որպես հիֆերի չտարբերվող զանգված։ Միայն համապատասխան պայմաններում ֆոտոբիոնտի հետ միանալիս առաջանում է բնութագրական ձևը։ Այդ պրոցեսը կոչվում է մորֆոգենեզ։ Հայտնի են դեպքեր, երբ միևնույն սունկը վերածվել է քարաքոսի երկու տարբեր ձևերի՝ միանալով մի դեպքում ջրիմուռի, մյուս դեպքում՝ ցիանոբակտերիայի հետ։ Սկզբում այդ երկու ձևերը համարվում էին երկու տարբեր տեսակներ, սակայն հետագայում հայտնաբերվեց, որ նրանք աճում են միացյալ կերպով։
Կան ապացույցներ, որ քարաքոսերում՝ բաղադիչների միջև, դիտվում է ոչ թե մուտուալիզմ, այլ կոմենսալիզմ կամ մակաբուծություն (Ահմաջյան,1993)։ Ֆոտոսինթեզող բաղադրիչը կարող է սնկից անկախ գոևատևել բնության մեջ, բայց հակառակը տեղի չի ունենում։ Ավելին՝ ֆոտոբիոնտի բջիջները պարբերաբար ոչնչացվում են սնկի կողմից սննդանյութերի փոխանակման ընթացքում։ Համակեցությունը պահպանվում է նրա շնորհիվ, որ ֆոտոբիոնտի բջիջները բազմանում են և լրացնում բջիջների պակասը։
Տիպիկ տերևանման քարաքոսի թալոմի կտրվածքը խոշորացման տակ ուսումնասիրելու ժամանակ պարզ դարձավ, որ սնկային ֆիլամենտների չորս շերտեր հաջորդում են միմյանց։ Ամենավերին շերտը կազմված է խիտ կերպով իրար կապակցված հիֆերից, որոնք առաջացնում են պաշտպանական արտաքին շերտ՝ կորտեքս (կեղև), որի հաստությունը կարող է հասնել մի քանի հարյուր միկրոմետրի։ Կորտեքսը կարող է լրացուցիչ կերպով ծածկված լինել բջջային կառուցվածք չունեցող շերտով՝ էպիկորտեքսով, որի հաստությունը մոտավորապես 0.6-1 մկմ է։
Ե´վ կանաչ ջրիմուռ, և´ ցիանոբակտերիա պարունակող քարաքոսերում ցիանոբակտերիաները գտնվում են վերին կամ ստորին մակերեսում գտնվող հատուկ փոքր կառույցներում՝ ցեֆալոդիումներում։ Վերին կորտեքսից ներքև գտնվում է ջրիմուռային շերտը՝ կազմված սնկի հիֆերի մեջ խիտ ձևով ներկառուցված ջրիմուռի բջիջներից։
Ֆոտոբիոնտի յուրաքանչյուր բջիջ կամ բջիջների խումբ սովորաբար փաթաթված է հիֆերով։ Որոշ դեպքերում այդտեղ ներթափանցում են սնկի հաուստորիումները։ Այս շերտի ներքևում գտնվում է երրորդ շերտը՝ կազմված թույլ կերպով միահյուսված սնկերի հիֆերից, այստեղ չկան ջիմուռների բջիջներ և այս շերտը կոչվում է մեդուլա։ Մեդուլայի տակ՝ ներքին մակերեսին, գտնվող շերտը նման է վերին շերտին և կոչվում է ստորին կորտեքս։ Այն վերին շերտի նման կազմված է խիտ ձևով փաթեթավորված հիֆերից։ Ստորին կորտեքսը կրում է ռիզիններ՝ արմատանման սնկային կառուցվածքներ, որոնք ծառայում են սուբստրատին ամրանալու համար։ Քարաքոսերը երբեմն պարունակում են նաև սնկային մետաբոլիտներից կազմված կառուցվածքներ։ Օրինակ՝ կեղևանման քարաքոսերը երբեմն կեղևում ունեն պոլիսախարիդային շերտ։ Չնայած յուրաքանչյուր քարաքոսի թալոմ հոմոգեն է թվում, կան որոշակի ապացույցներ, որ սունկը կազմված է տեսակի մեկից ավելի գենետիկական առանձնյակներից։ Նույնը կարծես ճիշտ է նաև ֆոտոբիոնտի տեսակների համար։
Աճման ձևեր[խմբագրել | խմբագրել կոդը]
Քարաքոսերին կարելի է դասակարգել ըստ իրենց աճման ձևերի։
- Կեղևանման, օրինակ՝ Caloplaca flavescens
- Թելանման, օրինակ՝ Ephebe lanata
- Տերևանման, օրինակ՝ Hypogymnia physodes
- Թփանման, օրինակ՝ Cladonia evansii, C. Subtenuis և Usnea australis
- Փոշենման, օրինակ՝ Lepraria incana
- Թեփուկանման, օրինակ՝ Normandina pulchella
- Դոնդողանման, որոնց մոտ ցիանոբակտերիաները արտադրում են ջուրը կլանող պոլիսախարիդ։
Բազմացումը և տարածումը[խմբագրել | խմբագրել կոդը]


Շատ քարաքոսեր բազմանում են անսեռ եղանակով։ Այն կատարվում է վեգետատիվ ճանապարհով կամ դիասպորների (պարունակում են սնկի և ջրիմուռի բջիջներ) տարածման միջոցով։ Դիասպորների տեսակներից են սոռեդիումները, որոնք ջրիմուռների բջիջների փոքրիկ խմբեր են։ Դրանք, շրջապատվելով սնկային ֆիլամենտներով, ձևավորում են հատուկ կառույցներ՝ սոռալիաներ, որոնց միջից սոռեդիումները կարող են տարածվել քամու միջոցով։ Դիասպորների մեկ այլ տեսակ են ինսիդիաները, որոնք թալոմի վրա գտնվող երկարավուն ելուստներ են և տարածվում են՝ պոկվելով թալոմից։ Այսպիսով, քարաքոսերն ունեն ինսիդիումներ, սոռեդիումներ և ենթարկվում են ֆրագմենտացիայի։ Չնայած այդ երկու կառուցվածքներն էլ կազմված են միևնույն բաղադրիչներից (միկոբիոնտ և ֆոտոբիոնտ), նրանք որոշակիորեն տարբերվում են։ Ինսիդիումները քարաքոսի արտաքին ելուստներ են։ Սոռեդիումները փոշենման պրոպագուլներ են, որոնք դուրս են գալիս թալոմի գագաթից։ Որպեսզի առաջանա քարաքոս, պրոպագուլները պետք է պարունակեն և´ միկոբիոնտ, և´ ֆոտոբիոնտ։
Թփանման քարաքոսերը հեշտորեն ֆրագմենտների են բաժանվում։ Քանի որ թալոմը լավ տարբերակված չէ, դիասպորների առաջացման և վեգետատիվ բազմացման միջև սահմանն անորոշ է։ Շատ քարաքոսեր կոտրվում են մասերի, երբ չորանում են և տարածվում են քամու միջոցով։ Շարունակում աճել, երբ առկա է անհրաժեշտ քանակի խոնավություն։
Շատ քարաքոսեր բազմանում են սեռական եղանակով՝ սնկերի նման առաջացնելով սպորներ մեյոզի արդյունքում։ Տարածվելուց հետո այս սպորները պետք է հանդիպեն համատեղելի ջրիմուռների հետ, որպեսզի առաջանա քարաքոս։ Բազմացման այս ձևը տարածված է բազիդիոլիխենների մոտ, որոնք ձևավորում են պտղամարմիններ։ Ասկոլիխենների մոտ սպորները ձևավորվում են հատուկ կառուցվածքներում՝ ապոտեցիումներում, պերիտեցիումներում և պիկնիդիումներում[11]։
Էկոլոգիա[խմբագրել | խմբագրել կոդը]
Քարաքոսերը մրցակցում են բույսերի հետ լույսի համար, սակայն իրենց փոքր չափսերի և դանդաղ աճի պատճառով լավ են աճում այն վայրերում, որտեղ բարձրակարգ բույսերը դժվարանում են աճել։ Քարաքոսերը միշտ առաջինն են բնակություն հաստատում այն վայրերում, որտեղ չկա հող, եզակի բուսականություն առաջացնում են մի շարք էքստրեմալ միջավայրերում, ինչպիսիք են բարձր լեռները, անապատները և հյուսիսային շրջանների սառցակալած հողերը։
Քարաքոսերի կարևոր էկոֆիզիոլոգիական առավելությունը նրանց պոյկիլոհիդրիկ լինելն է, ինչը նշանակում է, որ նրանք կարող են հանդուրժել անկանոն և երկարատև չորացումները։ Որոշ մամուռների, գետնամուշկերի և անապատային բույսերի նման քարաքոսերը ջրազրկման պայմաններում ևս դանդաղեցնում, կանգնեցնում են նյութափոխանակությունը (կրիպտոբիոզ), ինչի ժամանակ քարաքոսի բջիջները ջրազրկվում են այն աստիճան, որ ընդհատվում է կենսաքիմիական ակտիվությունը։ Կրիպտոբիոզի վիճակում քարաքոսերը ընդունակ են գոյատևել միջավայրի առավել դժվար պայմաններում՝ բարձր և ցածր ջերմաստիճաններում, ճառագայթման և չորացման պայմաններում։
Քարաքոսերը չունեն արմատներ և կարիք չունեն ջուր կլանել բարձրակարգ բույսերի նման, ուստի նրանք կարող են աճել այնպիսի վայրերում, որոնք անհասանելի են շատ բույսերի համար՝ մերկ ժայռերի վրա, անբերրի հողերի կամ ավազի վրա, ինչպես նաև արհեստական կառուցվածքների՝ պատերի, տանիքների և հուշարձանների վրա։ Շատ քարաքոսեր աճում են որպես էպիֆիտներ բույսերի վրա, մասնավորապես՝ ծառերի բների և ճյուղերի վրա։ Այս դեպքերում քարաքոսերը մակաբույծներ չեն, քանի որ բույսի վրա աճելու ժամանակ չեն սնվում բույսի որևէ մասով և չեն թունավորում այն։ Որոշ ցամաքային քարաքոսեր, ինչպես Cladina ենթացեղի ներկայացուցիչները արտադրում են ալելոպաթիկ նյութեր, որոնք ներծծվում են հողի մեջ և արգելակում բույսերի սերմերի ծլումը և երիտասարդ բույսերի աճը։ Սուբստրատի կայունությունը (երկարակեցությունը) քարաքոսերի բնակատեղի կարևոր գործոններից մեկն է։ Շատ քարաքոսեր աճում են կայուն ժայռային մակերեսներին կամ ծեր ծառերի կեղևների վրա, մյուս մասը՝ հողի և ավազի վրա։ Վերջին երկու դեպքերում քարաքոսերը կարևոր նշանակություն ունեն հողերի կայունացման հարցում. որոշ անապատային էկոհամակարգերում բարձրակարգ բույսերի սերմերը ծլում են միայն այն վայրերում, որտեղ քարաքոսերը կայունացնում են ավազը և օգնում կլանել ջուր։

Եվրոպական տիեզերական կազմակերպությունը բացահայտել է, որ քարաքոսերը կարող են գոյատևել տիեզերքում։ Լեոպոլդ Սանչոյի (Մադրիդի Կոմպլուտենսե համալսարան) կողմից կատարված փորձի ժամանակ Rhizocarpon geographicum և Xanthoria elegans տեսակի քարաքոսերը դրվել էին հատուկ կապսուլի մեջ և տեղադրվել ռուսական Սոյուզ հրթիռի մեջ՝ 2005 թվականի մայիսի 31-ին։ Արդեն տիեզերքում կապսուլը բացվել էր և քարաքոսերը հայտնվել էին տիեզերական տարածության մեջ, որտեղ առկա են ջերմաստիճանային խիստ տատանումներ և ճառագայթում։ 15 օր հետո քարաքոսերը ետ էին բերվել Երկիր, սակայն պարզվել էր, որ դրանք չունեին նկատելի վնասվածքներ և գտնվում էին առողջ վիճակում[12][13]։
Երբ քարաքոսերն աճում են հանքային մակերեսների վրա, որոշ քարաքոսեր քիմիական և ֆիզիկական եղանակներով դանդաղորեն քայքայում են սուբստրատները՝ առաջ բերելով հողմահարման գործընթացներ, որոնց արդյունքում ժայռերն աստիճանաբար վերածվում են հողի։ Չնայած այս գործընթացները հիմնականում օգտակար են, նրանք երբեմն կարող են խնդիրներ ստեղծել արհեստական քարե կառույցների համար։ Այսպիսի մի խնդիր է քարաքոսերի շարունակվող աճը Ռաշմոր լեռ ազգային հուշարձանի վրա, ինչը պահանջում է լեռնագնաց պահապանների միջամտություն՝ հուշարձանը մաքրելու համար։
Որոշ կենդանիներ սնվում են քարաքոսերով, օրինակ՝ հյուսիսային եղջերուն։ Թեփուկաթևավորների որոշ տեսակների թրթուրները սնվում են բացառապես քարաքոսերով։ Այնուամենայնիվ, քարաքոսերը պարունակում են շատ ածխաջրեր ու քիչ սպիտակուցներ և մի շարք կենդանիների համար անպիտան են։ Հյուսիսային թռչող սկյուռը օգտագործում է քարաքոսերը բույն սարքելու, սնվելու համար և որպես ջրի աղբյուր ձմռան ընթացքում։
Օդի աղտոտում[խմբագրել | խմբագրել կոդը]

Եթե քարաքոսերը միշտ աճում են աղտոտված օդի պայմաններում, նրանք ուղղակիորեն չեն կարող չկուտակել աղտոտիչները։ Նրանք չունեն թափվող մասեր, հերձանցներ, կուտիկալա և այդ պատճառով կլանում են տարբեր աերոզոլներ և գազեր թալոմի ամբողջ մակերեսով, որտեղից այդ նյութերը հեշտությամբ թափանցում են ֆոտոբիոնտի շերտի մեջ։ Արմատներ չունենալու պատճառով շատ քիմիական տարրերի առաջնային աղբյուրը քարաքոսերի համար օդն է, և այդ պատճառով քարաքոսի մեջ տարրերի պարունակությունը խոսում է նաև շրջակա օդի բաղադրության մասին։ Մթնոլորտից տարբեր նյութեր կլանվում են մառախուղի, ցողի, գազերի աբսորբցիայի և չոր նստեցման միջոցով։ Հետևաբար շատ էկոլոգիական ուսումնասիրություններ ընդծգում են քարաքոսերի շահութաբեր կիրառությունը որպես մթնոլորտային օդի որակի արդյունավետ կենսացուցիչներ։
Ոչ բոլոր քարաքոսերն են ցուցաբերում հավասար զգայունություն օդի աղտոտման նկատմամբ, այսինքն՝ քարաքոսերի տարբեր տեսակներ ունեն մթնոլորտի որոշակի աղտոտիչների նկատմամբ տարբեր զգայունություն։ Այդ զգայունությունը ուղղակիորեն կապված է միկոբիոնտի էներգետիկ պահանջների հետ, և որքան մեծ է միկոբիոնտի կախվածությունը ֆոտոբիոնտից, այնքան ավելի զգայուն է քարաքոսը օդի աղտոտվածության նկատմամբ։ Ֆոտոբիոնտը օգտագործում է նյութափոխանակային էներգիայի մի մասը բջջային կառուցվածքների վերանորոգման համար՝ ֆոտոսինթետիկ ակտիվության հաշվին, և այս դեպքում միկոբիոնտին բաժին է հասնում պակաս էներգիա։ Ֆոտոբիոնտի և միկոբիոնտի միջև հավասարակշռության խախտումը կարող է հանգեցնել սիմբիոտիկ համակեցության վերացմանը։ Հետևաբար քարաքոսի վերացումը կարող է տեղի ունենալ ոչ միայն թունավոր նյութերի կլանման, այլ նաև սիմբիոնտներից մեկի օգտին նյութերի անհավասարաչափ մատակարարման հետևանքով։
Էվոլյուցիա և հնէաբանություն[խմբագրել | խմբագրել կոդը]
Ասկոմիցետների և քարաքոսերի էվոլյուցիան բարդ է, և կան չպարզաբանված բազմաթիվ հարցեր, բայց քանի որ գոյություն ունեն ասկոմիցետների 15 տարբեր դասեր, գիտնականները հակված են հավատալ, որ տարբեր քարաքոսեր զարգացել են մեկը մյուսից անկախ՝ անալոգ էվոլյուցիայի ընթացքում։ Լիխենիզացված սնկերը շարունակել են զարգանալ քարաքոսեր չառաջացնող սնկերից անկախ։
Լիխենիզացումը սնկերի կողմից վաղուց ընտրված մարտավարություն է։ Էքստրեմալ բնակավայրերը, որտեղ քարաքոսերը ապրում են, սովորաբար չեն նպաստում բրածոների առաջացմանը։ Քարաքոսերի ամենահին բրածոն, որում երկու սիմբիոտիկ բաղադրիչներն էլ պահպանվել են, պատկանում է վաղ Դեվոնի ժամանակաշրջանին և ունի 400 միլիոն տարվա վաղեմություն[14]։ Մի փոքր ավելի հին բրածո է Սպոնգիոֆիտոնը, որը նույնպես ներկայացվում է որպես քարաքոս։ Կա դեռևս չապացուցված ենթադրություն, որ ավելի հին բրածո Նեմատոթալուսը նույնպես քարաքոս է։
Կան պնդումներ, որ Էդիակարյան բրածոները ևս քարաքոսեր են չնայած կան որոշ հակասություններ՝ կապված նրանց թվին պատկանող Դիկինսոնիաի հետ։ Հարավային Չինաստանում՝ Դոուշանտյո ֆորմացիայի ծովային ֆոսֆորիտներում, պահպանվել են քարաքոսանման բրածոներ՝ կազմված կլորավուն բջիջներից և բարակ ֆիլամենտներից։ Այս բրածոները պատկանում են Նեոպրոտերոզոիկ դարաշրջանին և ենթադրաբար ունեն 551-635 միլիոն տարվա վաղեմություն[15]։ Այս բրածոների բացահայտումը խոսում է այն վարկածի օգտին, որ սնկերը ձևավորել են սիմբիոտիկ հարաբերություններ ֆոտոավտոտրոֆների հետ բարձրակարգ բույսերի էվոլյուցիայից շատ առաջ։ Վինֆրենատիան, որը գտնվել է Շոտլանդիայում, նույնպես վերագրվում է վաղ դեվոնյան ժամանակներին և իրենից ներկայացնում է զիգոմիցետային քարաքոս։ Գտնվել են մի շարք բրածո քարաքոսեր՝ սաթի մեջ պարփակված վիճակում։ Օրինակ՝ Anzia անունով բրածո քարաքոսը հայտնաբերվել է սաթի կտորների մեջ՝ հյուսիսային Եվրոպայում և ունի մոտ 40 միլիոն տարվա վաղեմություն։ ԱՄՆ-ի Կալիֆոռնիա նահանգի Թրինիթիի շրջանում հայտնաբերված Lobaria-ն պատկանում է միջին միոցենի ժամանակաշրջանին։
1995 թվականին Գարգասը և նրա գործընկերները առաջարկեցին մի վարկած, որ գոյություն ունի լիխենիզացման ամենաքիչը 5 անկախ աղբյուր՝ 3-ը բազիդիոմիցետներում և 2-ը ասկոմիցետներում։ Այնուամենայնիվ Լուցոնին և մի շարք այլ գիտնականներ 2000 թվականին նշեցին, որ լիխենիզացումը հավանաբար զարգացել է ավելի վաղ և ուղեկցվել է բազմակի կորուստներով։ Գուցե քարաքոս չառաջացնող որոշ սնկեր երկրորդաբար կորցրել են քարաքոսային ասոցիացիա առաջացնելու ընդունակությունը։ Արդյունքում, լիխենիզացումը համարվեց որպես սննդառության առումով հաջողված մարտավարություն։
Քարաքոսերը եղել են հին ցամաքային էկոհամակարգերի բաղադրիչներ։ Ամենահին բրածոն ենթադրվում է, որ ունի 400 միլիոն տարվա տարիք։ Վերջին հետազոտությունները (2009) ցույց են տվել, որ ասկոմիցետների նախնիները եղել են սապրոֆիտներ, և որ լիխենիզացման պրոցեսները տեղի են ունեցել բազմակի անգամներ։
Կարգաբանություն և դասակարգում[խմբագրել | խմբագրել կոդը]
Քարաքոսերը անվանվում են սնկային բաղադրիչի հիման վրա, քանի որ այն որոշիչ դեր է խաղում քարաքոսի ձևի որոշման հարցում։ Սունկը հիմնականում կազմում է քարաքոսի մարմնի հիմնական մասը, չնայած թելանման և դոնդողանման քարաքոսերի դեպքում այդպես չէ։ Սունկը հիմնականում ասկոմիցետ է, քիչ դեպքերում՝ բազիդիոմիցետ, եվ համապատասխանաբար տարբերում ենք բազիդիոլիխեններ և ասկոլիխեններ։ Նախկինում որոշ կարգաբաններ մտցնում էին քարաքոսերին առանձին բաժնի մեջ, որը կոչվում էր Mycophycophyta, սակայն այս դասակարգումը այլևս չի ընդունվում, քանի որ քարաքոսի բաղադրիչները ունեն տարբեր ծագում։ Ո´չ ասկոլիխենները և ո´չ էլ բազիդիոլիխենները չեն առաջացնում մոնոֆիլետիկ սերունդներ, բայց առաջացնում են որոշակի խմբեր։ Յուրահատուկ սունկ է Geosiphon pyriforme-ն, որը պատկանում է Glomeromycota տիպին և ներառում է ցիանոբակտերիալ սիմբիոնտին իր բջիջների մեջ։
Geosiphon-ը
սովորաբար չի համարվում քարաքոս, և իր յուրահատուկ սիմբիոզը չէր ընդունվում տարիներ շարունակ։ Այս ցեղը ավելի մոտ է կանգնած էնդոմիկորիզա առաջացնող սնկերի ցեղերին։
Հետևալ աղյուսակում տրված են քարաքոս առաջացնող սնկերի կարգերը և ընտանիքները տես ստորև։
Տնտեսական նշանակություն[խմբագրել | խմբագրել կոդը]

Քարաքոսերն օգտագործվում են որպես սնունդ աշխարհի տարբեր ժողովուրդների կողմից։ Չնայած որոշ քարաքոսերով սնվում են միայն սովի ժամանակ, մյուս տեսակները օգտագործվում են որպես հիմնական սնունդ և նույնիսկ որպես դելիկատես։ Քարաքոսերով սնվելու ժամանակ առաջանում են որոշակի դժվարություններ. քարաքոսի պոլիսախարիդները անմարսելի են մարդկանց համար, ինչպես նաև սովորաբար պարունակում են թույլ թունավոր երկրորդային մետաբոլիտներ, որոնք պետք է հեռացվեն ուտելուց առաջ։ Շատ քիչ քարաքոսեր են թունավոր, որոնք հիմնականում դեղին են։
Իսլանդական մամուռ կոչվող քարաքոսը (Cetraria islandica) անցյալում Հյուսիսային Եվրոպայի բնակիչների սննդի անբաժան մասն էր կազմում։ Նրանից պատրաստում էին հաց, շիլա, պուդինգ, ապուր և աղցան։ Ֆրեմոնտի բրիորիա կոչվող քարաքոսը (Bryoria fremontii) լայնորեն օգտագործվում էր սննդի մեջ Հյուսիսային Ամերիկայում։ Հյուսիսային ժողովուրդները օգտագործում էին մասնակիորեն մարսման ենթարկված եղջերուի մամուռ կոչվող քարաքոսը՝ (Cladina spp.) այն նախապես հանելով սպանված եղջերուի ստամոքսի գանձակից։ Umbilicaria spp. և Lasalia spp. քարաքոսերը հաճախ օգտագործվել են Հյուսիսային Ամերկայում որպես սնունդ ծայրահեղ դեպքերում, իսկ Umbilicaria esculenta տեսակը կորեական և ճապոնական խոհանոցում է օգտագործվում։
Շատ քարաքոսեր արտադրում են երկրորդային մետաբոլիտներ՝ պիգմենտներ և տոքսիններ։ Պիգմենտները պաշտպանում են քարաքոսին արեգակնային հավելյալ ճառագայթումից, իսկ տոքսինները ոչնչացնում են բակտերիաներին և պաշտպանում են խոտակեր կենդանիներից։ Այս նյութերը կարևոր նշանակություն ունեն քարաքոսերի տեսակային պատկանելիությունը որոշելու ժամանակ և ունեն տնտեսական նշանակություն, որպես ներկերի (օրինակ՝ լակմուսի) և պարզ հակաբիոտիկների ստացման աղբյուր։
Կան տեղեկություններ, որ քարաքոսերը օգտագործվել են կարմիր և մանուշակագույն ներկեր ստանալու համար։ Կարևոր պատմական, տնտեսական նշանակություն ունեն Roccellaceae ընտանիքի քարաքոսերը, որոնք հաճախ կոչվում են օրխելլայի սերմեր կամ օրխիլներ։ Նրանցից ստացվող օրսեինը և այլ քարաքոսային ներկեր հետագայում փոխարինվեցին սինթետիկ տարբերակներով։ pH-ի ինդիկատոր լակմուսը ներկ է՝ ստացված Rocella tinctoria քարաքոսից եռացման միջոցով։
Usnea ցեղի տարբեր տեսակներ օգտագործվել են վերքերը բուժելու համար 20-րդ դարի կեսերին Ռուսաստանում։ Պատմականորեն Lobaria pulmonaria տեսակը հավաքվում էր մեծ քանակություններով և, քանի որ նման էր արտաքին տեսքով թոքերին, ապա վաճառվում էր որպես թոքային հիվանդությունների դեմ դեղ։ Նույն կերպով Peltigera leucophlebia տեսակը առաջարկվում էր օգտագործել կաթնուկի ժամանակ։
Օլիվետոլ կոչվող նյութը առկա է որոշ քարաքոսերի կազմում։ Կանեփի բույսը նույնպես արտադրում է նման մի նյութ՝ օլիվետոլաթթու՝ որպես միջանկյալ պրոդուկտ։ Այն հետագայում օգտագործվում է տետրահիդրոկանաբինոլի (THC) կենսասինթեզի ժամանակ։
PLoS ONE ամսագրում տպագրված մի հոդվածում նշված է, որ որոշ տեսակի քարաքոսեր քայքայում են պրիոններին։
Պատկերասրահ[խմբագրել | խմբագրել կոդը]
-
 Տերևանման քարաքոս Xanthoparmelia cf. lavicola-ը՝ բազալտի վրա
Տերևանման քարաքոս Xanthoparmelia cf. lavicola-ը՝ բազալտի վրա -
 Թփանման քարաքոս Usnea australis-ը՝ ծառի ճյուղի վրա
Թփանման քարաքոս Usnea australis-ը՝ ծառի ճյուղի վրա -
 Քարտեզ քարաքոս (Rhizocarpon geographicum) քարի վրա
Քարտեզ քարաքոս (Rhizocarpon geographicum) քարի վրա -
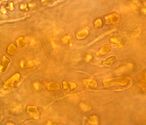 Ցիանոբակտերիա Hyella caespitosa-ն սնկային հիֆերով՝ Pyrenocollema halodytes քարաքոսի մոտ
Ցիանոբակտերիա Hyella caespitosa-ն սնկային հիֆերով՝ Pyrenocollema halodytes քարաքոսի մոտ -
 Ցիանոբակտերիա Hyella caespitosa-ն սնկային հիֆերով՝ Pyrenocollema halodytes քարաքոսի մոտ
Ցիանոբակտերիա Hyella caespitosa-ն սնկային հիֆերով՝ Pyrenocollema halodytes քարաքոսի մոտ -
 Cladonia rangiferina՝ եղջերուի քարաքոս
Cladonia rangiferina՝ եղջերուի քարաքոս -
 Hypogymnia cf. tubulosa, Bryoria sp. և Tuckermannopsis sp. Կանադական ժայռոտ լեռներ
Hypogymnia cf. tubulosa, Bryoria sp. և Tuckermannopsis sp. Կանադական ժայռոտ լեռներ -
 Lobaria oregano, Հոհ անտառ, Վաշինգտոն
Lobaria oregano, Հոհ անտառ, Վաշինգտոն








Արտաքին հղումներ[խմբագրել | խմբագրել կոդը]
- Հայաստանի քարաքոսեր Արխիվացված 2021-02-04 Wayback Machine
- Դպրոցական Մեծ Հանրագիտարանում
- Քարաքոսերի մասին դասարան․ամ կայքում Արխիվացված 2016-09-06 Wayback Machine
Գրականություն[խմբագրել | խմբագրել կոդը]
Հայերեն[խմբագրել | խմբագրել կոդը]
- Փանոսյան, Հ. Կ. and Նիկողոսյան, Վ. Գ. (1973) Հայաստանի քարերի վրա զարգացող ամենատարածված քարաքոսների մասին. Հայաստանի կենսաբանական հանդես Biological Journal of Armenia Биологический журнал Армении ՀՍՍՀ ԳԱ, 26 (10). pp. 3–9. ISSN 0366-5119
- Harutyunyan, S., Wiesmair, B. & Mayrhofer, H. 2011. Catalogue of the lichenized fungi in Armenia. Herzogia 24։ 265 –296
- Gasparyan A. & Sipman H. J. M. 2013. New lichen records from Armenia // Mycotaxon, vol 123: 491 (link page).
- Gasparyan A. H. & Ghaltaghchyan Ts. K. 2013. Proposed standard designations in Armenian for lichens occurring in the Republic of Armenia // Fl. Medit., 23: 105-116.
- Gasparyan A., Sipman H. J. M. & Brackel W. von. 2014. A contribution to the lichen-forming and lichenicolous fungi flora of Armenia. – Willdenowia 44: 263 – 267.
Անգլերեն[խմբագրել | խմբագրել կոդը]
- Ahmadjian V. (1993). The Lichen Symbiosis. New York։ John Wiley & Sons. ISBN 0-471-57885-1.
- Brodo, I.M., S.D. Sharnoff, and S. Sharnoff, 2001. Lichens of North America. Yale University Press, New Haven.
- Gilbert, O. 2004. The Lichen Hunters. The Book Guild Ltd. England.
- Haugan, Reidar; Timdal, Einar (1992). "Squamarina scopulorum (Lecanoraceae), a new lichen species from Norway". Nordic Journal of Botany 12 (3)։ 357–360. doi։10.1111/j.1756-1051.1992.tb01314.x.
- Hawksworth, D.L. and Seaward, M.R.D. 1977. Lichenology in the British Isles 1568 - 1975. The Richmond Publishing Co. Ltd., 1977.
- Kershaw, K.A. Physiological Ecology of Lichens, 1985. Cambridge University Press Cambridge.
- Kirk PM, Cannon PF, Minter DW, Stalpers JA. (2008). Dictionary of the Fungi. (10th ed.). Wallingford։ CABI. ISBN 978-0-85199-826-8.
- Knowles M.C. (1929). "Lichens of Ireland". Proceedings of the Royal Irish Academy 38։ 1–32.
- Purvis, O.W., Coppins, B.J., Hawksworth, D.L., James, P.W. and Moore, D.M. (Editors) 1992. The Lichen Flora of Great * Britain and Ireland. Natural History Museum, London.
- Sanders W.B. (2001). "Lichens: interface between mycology and plant morphology". BioScience 51 (12)։ 1025–1035. doi։10.1641/0006-3568(2001)051[1025։LTIBMA]2.0.CO;2.
- Seaward M.R.D. (1984). "Census Catalogue of Irish Lichens". Glasra 8։ 1–32.
- Whelan, P. 2011. Lichens of Ireland. The Collins Press, Cork, Ireland.
Ռուսերեն[խմբագրել | խմբագրել կոդը]
- А. Н. Окснер. Определитель лишайников СССР. Вып. 2. Морфология, систематика и географическое распространение / отв. ред. И. И. Абрамов. - Л.։ «Наука», 1974.
- Определитель лишайников СССР. Л., 1975. Вып. 3. С. 85 - 105.
- Водоросли, лишайники и мохообразные СССР / отв. ред. М. В. Горленко. - М.։ «Мысль», 1978.
- Лишайники // 2003 * Россия * Красный список особо охраняемых редких и находящихся под угрозой исчезновения животных и растений / ВНИИ охраны природы, Лаборатория Красной книги; Отв. ред. В. Е. Присяжнюк. М., 2004. Вып. 2, ч. 4։ Споровые растения и грибы. С. 189 - 250.
Ծանոթագրություններ[խմբագրել | խմբագրել կոդը]
- ↑ Cambridge Advanced Learner's Dictionary Second Edition, page 731. Cambridge University Press, 2005
- ↑ F.S. Dobson (2000) Lichens, an illustrated guide to the British and Irish species. Richmond Publishing Co. Ltd., Slough, UK
- ↑ Morris J, Purvis W. (2007). Lichens (Life). London: The Natural History Museum. էջ 19. ISBN 0-565-09153-0.
- ↑ Ferry, B.W., Baddeley, M.S. & Hawkworth, D. L. (Editors) (1973) Air Pollution and Lichens. Athlone Press, London.
- ↑ Rose C.I., Hawksworth D.L. (1981). «Lichen recolonization in London's cleaner air». Nature. 289 (5795): 289–292. Bibcode:1981Natur.289..289R. doi:10.1038/289289a0.
- ↑ Baldwin, Emily (2012 թ․ ապրիլի 26). «Lichen survives harsh Mars environment». Skymania News. Արխիվացված է օրիգինալից 2012 թ․ մայիսի 28-ին. Վերցված է 2012 թ․ ապրիլի 27-ին.
- ↑ de Vera, J.-P.; Kohler, Ulrich (2012 թ․ ապրիլի 26). «The adaptation potential of extremophiles to Martian surface conditions and its implication for the habitability of Mars» (PDF). European Geosciences Union. Արխիվացված է օրիգինալից (PDF) 2012 թ․ հունիսի 8-ին. Վերցված է 2012 թ․ ապրիլի 27-ին.
- ↑ Heribert Schöller: Flechten — Geschichte, Biologie, Systematik, Ökologie, Naturschutz und kulturelle Bedeutung. Schweizerbartsche Verlagsbuchhandlung, Frankfurt am Main 1997, ISBN 3-7829-1151-2, S. 3-10
- ↑ Honegger R. (2000). «Simon Schwender (1829–1919) and the dual hypothesis in lichens». Bryologist. 103 (2): 307–13. doi:10.1639/0007-2745(2000)103[0307:SSATDH]2.0.CO;2. ISSN 0007-2745. JSTOR 3244159.
- ↑ Rikkinen J. (1995). «What's behind the pretty colors? A study on the photobiology of lichens». Bryobrothera. 4: 1–226.
- ↑ «BBC NEWS | UK | Scotland | Insight into sex life of lichens». BBC News. 2006 թ․ սեպտեմբերի 18. Վերցված է 2010 թ․ փետրվարի 16-ին.
- ↑ «ESA — Human Spaceflight and Exploration - Lichen survives in space». Վերցված է 2010 թ․ փետրվարի 16-ին.
- ↑ Sancho, L.G.; De La Torre, R.; Horneck, G.; Ascaso, C.; De Los Rios, A.; Pintado, A.; Wierzchos, J.; Schuster, M. (2007). «Lichens survive in space: results from the 2005 LICHENS experiment». Astrobiology. 7 (3): 443–54. Bibcode:2007AsBio...7..443S. doi:10.1089/ast.2006.0046. PMID 17630840.
{{cite journal}}: CS1 սպաս․ բազմաթիվ անուններ: authors list (link) - ↑ Taylor, T.N.; Hass, H.; Remy, W.; Kerp, H. (1995). «The oldest fossil lichen». Nature. 378 (6554): 244–244. doi:10.1038/378244a0.
{{cite journal}}: CS1 սպաս․ բազմաթիվ անուններ: authors list (link) - ↑ Yuan X, Xiao S, Taylor TN. (2005). «Lichen-like symbiosis 600 million years ago». Science. 308 (5724): 1017–20. Bibcode:2005Sci...308.1017Y. doi:10.1126/science.1111347. PMID 15890881.
{{cite journal}}: CS1 սպաս․ բազմաթիվ անուններ: authors list (link)
 |
Այս հոդվածն ընտրվել է Հայերեն Վիքիպեդիայի՝ {{{1}}} թվականի {{{2}}} շաբաթվա հոդված։ |